|
Over the last several years I have spent
a considerable amount of time trying to convince marine reef
aquarists that one of the most useful and important components
of a coral reef aquarium system is the deep sand bed. With
over thirty years of experience as a marine ecologist studying
the interactions in sediment ecosystems, I have acquired a
tremendous urge to discuss the importance of such beds in
great and wondrous detail, and I have been fortunate enough
to have been able to do so at numerous conferences and aquarium
societies. Often, however, this has had the interesting side
effect of causing most people to drift gently into dreamland.
While such events are indeed a cause for puzzlement, I have
decided that exploring its cause would be less than enlightening.
Rather in this month's column, I decided to forego my usual
quarterly discussion of reef aquarium sand beds.
Instead, I thought I would take this opportunity
to acquaint you with some of the most unusual organisms that
make their way into coral reef aquaria, the Foraminifera.
Forams, as they are commonly called, are marine creatures
found in a wide variety of habitats. They are not animals;
they lack not only a number of animal characteristics, but
also the photosynthetic capabilities of organisms such as
plants or algae. So, biologists consider they are neither
plants nor animals, and they are taxonomically placed in a
separate kingdom, with a number of other groups of bizarre
creatures. This kingdom is called the Protista. Even though
they are not animals, some protists, such as forams and some
other protozoans, act somewhat like animals.
While many protozoans are not clearly visible
to the unaided eye, forams are often evident and common in
many marine environments, and those found in aquaria are often
large enough to be observed easily with a hand lens or magnifying
glass. Even so, though, they tend not to show too much detail.
This is due to two reasons, first the detail that is there
is often very tiny, below the range of normal visual acuity,
and secondly, due in part to the type of body these organisms
have there is often not much detail to see. They simply don't
have a body divided into separate parts or structures to see
any detail.
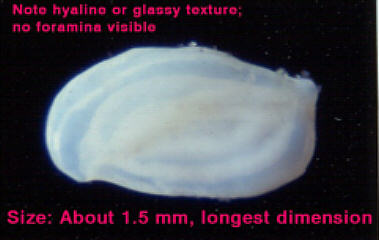
Figure
1. A foraminiferan collected from my aquarium.
Many of the forams found in aquaria are
best observed with a hand lens. These are available on-line
from biological or geological supply houses and will magnify
ten to twenty times. Using such a lens, an aquarist will be
able to clearly see foraminiferans and many other small organisms
if they are near the aquarium walls. Magnifying glasses will
also work, but they typically don't magnify the organisms
sufficiently.
As the typical aquarist discovers, our
hobby is rife with unusual organisms. Coral reef organisms,
in general, are not that familiar to most folks. We are terrestrial
animals, and the other animals we are familiar with are also
terrestrial organisms. Oceanic environments, such as coral
reefs and the deep sea contain strange and often wonderful
creatures. Our aquaria are proof of that, but even amongst
the strange and wonderful organisms of coral reefs, foraminiferans
classify as a "wee bit weirder" yet. Foraminiferans
are truly amongst the most unusual creatures in our systems.
Forams and their relatives are organisms
that are simply constructed; they may be described as an amoeba
in a shell. Probably the most basic and primitive type of
animal-like living creatures that many people ever see are
the amoebas often used in introductory biology or even a general
science course to show a "primitive" single cell.
However, that so-called primitive characteristic is misleading.
Even a common amoeba is many thousands of times larger and
more complex than a bacterium, an organism that is a much
more basic and primitive form of life. To some extent, then,
the apparent simplicity of an amoeba or foraminiferan is misleading.
On the other hand, the level of intracellular structure seen
in these organisms is quite low compared to such complicated
protists as ciliates.
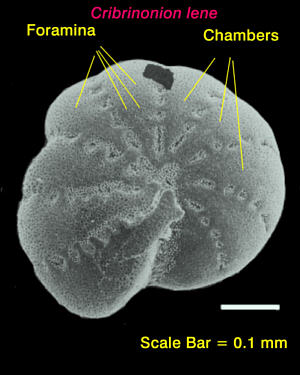 |
Figure
2. Scanning electron micrograph of a test or shell
of
the foraminiferan, Cribrononion lene. The small pores,
or "foramina," found in the test or shell are the
basis of the
name for the group of organisms.
The amoeba that most people have seen is
the common Amoeba proteus or the delightfully named
Chaos chaos. However, these particular species are
only a couple of the many kinds of related organisms at this
level of body structure. Many people consider that amoebas
and other protozoans are organisms that are made of only one
cell. Another, equally valid way of looking at them, however,
is as organisms without any cells at all. How we consider
these organisms, as having one cell or being without cells,
significantly colors our appreciation of them. We are organisms
constructed of many trillions of cells, and we tend to, with
more than a little bit of hubris, look upon ourselves as the
pinnacle of life. From this point of view, a lowly little
one-celled blob is hardly worth any consideration at all.
On the other hand, if we consider both ourselves and amoebas
simply as organisms, we can see that these little blobs are
more than they might otherwise seem, for both humans and amoebas
are organisms, and must face some conceptually similar problems.
All organisms must meet and successfully pass certain tests.
They must obtain nutrition, grow, reproduce, sense the environment,
and avoid being eaten. It may well be that amoebas are primitive.
It may also be that they are very well adapted to a wholly
different environment than we are, and are in fact, within
that context not any less successful than any other widespread
organism.
Nonetheless, the amoebas are not large
complex multicellular animals. They are minute blobs of protoplasmic
goo that are adapted to slither around on wet surfaces, either
under water, in wet soil, or inside of other organisms. They
have no defined shape or orientation. Front is simply the
direction they are moving at the moment. Their body is almost
infinitely deformable. They are small enough that they have
no need for any special system to eliminate wastes, send nervous
impulses or eat. They eat by simply enfolding their body surface
to enclose a tasty food item in a bubble of cell membrane
material. Once encased inside their body, digestive enzymes
are secreted into the bubble surrounding the food item and
it is dissolved. Indigestible food is simply expelled from
the body. Respiration or other gas exchange simply occurs
across their body surface.
Foraminiferans have been called the "the
most common group of non-bacterial organisms in the world."
They are in uncountable numbers in and on all ocean bottoms
and in the marine plankton. I have worked with some foraminiferan
predators called scaphopods. In some of the areas where the
scaphopods are abundant and I have sampled foraminiferans,
there are more than 70,000 forams per square yard of ocean
bottom. These abundances appear to be typical or even on the
low side for some marine environments. In some places, skeletons
from dead planktonic foraminiferans have settled to the ocean
bottoms to form layers of "foraminiferan ooze" more
than 6,600 feet thick. They are also quite abundant in most
natural coral reefs and in reef aquarium systems, yet most
aquarists remain unfamiliar with them.
Here is an image
of the test or shell of the pelagic foram Globigerina
sp.
Many fantastic images of forams, living
and dead, can be found by following this link.
Although foraminifera are often thought
of as simply being amoebas possessing an outside shell, there
is more to them than that. Not the least of the problems with
such a simplistic approach is that the shell is actually interior
to the outer cell membrane that constitutes the outer surface
of the organism. Nonetheless, these shells act as support
and protection for the majority of the protoplasm that constitutes
the foram's body. Three basic shelled foram types may be recognized,
defined on the basis of differences in their shell, and recently
a few naked forams have been found as well. The naked forams
are unusual for a second reason, that being they are found
in fresh water. All other forams are marine. Foraminifera
with the first skeletal type are called agglutinated or arenaceous
forams. They glue sand and other materials together to form
an irregular, often star- or tree- shaped structure. These
organisms are very common in some coral reefs, particularly
in areas where sponges are common. In these areas they may
form "spicule trees;" three-dimensional structures
made of sponge spicules glued together extending up into the
water column. However, these irregular, or agglutinated, forams
really come into dominance in the deep seas. Here some species
may get large, about the size of dinner plates, perhaps larger.
Some species extend up off the bottom as a tree-like shape
and have been documented to snare and eat fish or small shrimps.
Others form networks of root-like growths that may cover large
areas. None of this group has, to the best of my knowledge,
been seen in aquaria, and in fact, only a few individuals
of these deep- sea groups have ever been seen alive.
Here are links to some pictures of arborescent
forams from Antarctica plus information on their biology.
http://scilib.ucsd.edu/sio/nsf/fguide/protoctista10.html
http://scilib.ucsd.edu/sio/nsf/gallery/sam6.jpg
Another type of foraminiferan secretes
its shell wholly out of organic materials, chiefly protein.
One common variety of this group looks like shiny brown spheres,
about one-twentieth of an inch or so in diameter. In nature,
they move slowly up and down algal stalks or across the substrate
collecting and eating particulate material. Their food is
mostly bacteria and microalgae. I have seen a few of these
in some aquaria, and they may actually be pretty common in
some hobbyist's tanks, but their small size and unspectacular
color tends to camouflage them.
The final kind of shelled foram secretes
a shell, totally or mostly, out of calcium carbonate. These
shells may be spherical, discoidal, tubular, or some other
odd shapes. They range in size from about one tenth of a millimeter
in diameter to giants, referred to as mermaid's pennies, well
over a inch in diameter. They are very abundant and quite
common around reefs tropics as well as in our aquaria. These
organisms move very slowly. Extended around their bodies are
meshes of fine strands of protoplasm referred to as "reticulopodia."
They use these to ingest the bacteria and other small organic
particulate material that is their main food.
A beautiful photo of a calcareous test
may be found by following this link.
Here is a link to some calcareous forams
living on a limpet in Antarctica.
Here is a link
to a tropical foram with its feeding filaments extended. This
species is common in reef aquaria.
Shelless or "naked" fresh-water
(!) foraminiferans have recently been described, primarily
on the basic of body form, the presence of reticulopodia,
and cellular chemistry. Very little is known about their biology.
Some information about the fresh water
forms, including an image, may be found by following this
link.
In Indo-Pacific coral reef areas, the calcareous-shelled
forams may be exceptionally abundant. In fact, the beach sands
around some atolls, such as some of the islets in Palau, may
be almost entirely made from foram shells. Such abundance
is not limited to the Pacific; the pink sands of Bermuda get
their color from pink foram skeletons; primarily from Homotrema
rubrum, a foram that grows abundantly in some aquaria.
It should not be surprising then, that foraminiferans may
become common in our systems. Presumably they enter our systems
on live rock, live sand, or coral and thrive.
Cells are considered to be the basic unit
of life. Some cells, those found in bacteria and archaeans
(a form of life that is somewhat similar to bacteria) are
very tiny and lack much in the way of visually evident internal
structure. The cells of animals, plants, fungi and most protists
are much larger and complex, and contain a lot of microscopically
visible intracellular machinery referred to as organelles.
The one type of organelle found in almost all larger cells
is the nucleus. This is the site of "cellular control"
and contains the genetic material that defines the cell and
cellular functions. If people think of cells at all, or if
they think of the cells that larger organisms are composed
of, they generally assume that most cells have only one nucleus.
Most cell types in multicellular animals, indeed, do have
only one nucleus. In mammals, some exceptions are striated
(or skeletal) muscle cells which have many nuclei, and mature
red blood cells which don't have a nucleus.
One feature that sets foraminiferans apart
from many other protozoans is the fact that they may be multinucleate.
Smaller forams often have one nucleus, but larger forams may
have several thousand nuclei within their body. Nevertheless,
forams do not have bodies divided up into cells. The lack
of discrete cells is one reason they are not considered to
be animals. In the large agglutinated forams, the nuclei seem
to be more-or-less distributed throughout the protoplasmic
mass. In the shelled forams, the main mass of the body, including
most of the nuclei, is found inside the shell, but a large
amount extends outward into or over the surrounding substrate
in a mesh-like network of fine fibers, the reticulopodia.
These fibers are their site of food collection. An image of
a giant foram from Antarctica showing feeding pseudopods,
plus lots of neat information may be found by following this
link.
Most forams are presumed to eat bacteria,
other protozoans, or fine particulate material by ingesting
them when they come in contact with the fiber network. However,
the larger forams are predators on just about anything they
can catch. A large foraminiferan, with a shell about a tenth
of an inch in diameter, may have fibers that extend outwards
over a half an inch from the shell. Food items are ingested
and eaten, and as the foram clears an area of acceptable food,
it slowly moves across or through the sediments.
In a deep sand bed, feeding foraminiferans
perform several vital functions for the aquarist. First, they
clear small particulate material from between the sediment
grains, allowing water movement, preventing stagnation. Second,
such feeding opens space on individual sediment grains, in
a manner analogous to forest fires opening spots in a forest.
This allows more bacterial population growth to occur, and
as the metabolism leading to this growth IS the biological
filter, this clearing of space is necessary for continual
biological filtration. Finally, when the forams metabolize
their ingested material, soluble mineral nutrients such as
phosphate and ammonium, ions are formed and excreted as waste.
In turn, these materials may be utilized by algae and converted
into a form that may be exported from the system simply by
subsequent harvesting of the algae. So, foraminiferans are
a vital link in the process leading to the removal of excess
nutrients caused by the necessity of feeding all the animals
in the system.
Some of the most abundant forams that are
found in our aquaria are spectacular. They are large and obvious
and are often exceptionally abundant. And, unlike the remainder
of the forams discussed in this article, they are not found
in the sediments. As they don't look like anything else in
our systems, they are often misidentified as small sponges,
hydrocorals or stony corals. These foraminiferans belong to
a peculiar foram species called Homotrema rubrum and
it has a shell that may be orange, but is more typically hot
pink or bright cherry red. The red coloration is due to an
iron salt that is incorporated into the skeleton. Found growing
on hard surfaces such as rocks, the calcareous shell looks
like a small hydrocoral or a hard, spiky crystal with angular
projections. Homotrema seldom get larger than an eighth
or quarter of an inch in height, but their brilliant color
renders them very obvious. They feed on particulate material
in the tank's water, probably mostly bacterial aggregates
they catch in fine filamentous protoplasmic strands which
extend from the tips of the angular projections. There are
similar white foraminiferan species found in aquaria. Their
bodies look like small "spiky" versions of a stony
coral, rather like nano-sized versions of bird's nest coral,
Seriatopora hystrix. These foraminiferans have not
been identified to species.
 |
Figure
3. A Homotrema rubrum; about one-third of an inch
high.
The rhizopodia are feeding structures.
The other forams found in our aquaria are
less evident, but more typical in shape. Some individuals
may actually be about as large as Homotrema and they
are often far more abundant. In some aquaria, these are actually
the most abundant "animal-like" organisms. And they
generally are completely overlooked. They are usually colored
some shade of white, beige or tan, although some common species
are pink. These species are typically spherical, ovoid, or
discoidal and can't be identified to species without specialized
references. Sometimes they may be seen crawling on rocks,
algae, or tank walls, but most of the time they are found
in the sediments. Many of them are about the size of a sesame
seed.
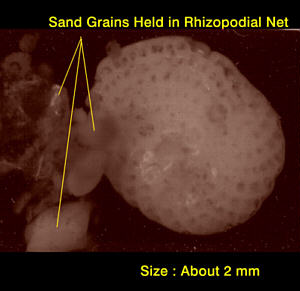
Figure
4. A large aquarium foram attached to sand grains by
its
rhizopodial feeding network.
An easy way to see if your system has an
abundant foram population is to sample a bit of the sand bed.
Remove a small amount of sediment from the surface of your
sand bed using a turkey baster or some other implement. Put
it into a clean clear glass bowl or dish, making sure it is
covered with water. You don't need much sediment for this
evaluation, just enough to make a layer one sand grain deep,
and there should be plenty of open area on the bottom of the
bowl. Then examine the sediments with a hand lens or magnifying
glass, and look for clumps of sand grains that are attached
to other sand grains by "mucous" masses. On close
examination, many of these masses will be seen to have a large
discoidal or spherical "grain" in them. This will
likely be a foram.
Figure
5. Some forams collected in about 10 minutes from one
of my aquaria.
It takes microscopic examination of the
sediments to confirm the presence of foraminiferans. But if
you are one of those hobbyists with access to a microscope,
you should examine the "suspect" sand grain for
the appearance of fine dots on the surface. The name Foraminifera
is derived from a combination of Latin and Greek terms meaning
"bearing pores or holes" and the surface of most
foram shells are covered with microscopic holes visible at
about 40x magnifications. If you are lucky, you may find larger
greenish or grayish-green foraminiferans about a quarter of
an inch in diameter. These are forams that have zooxanthellae.
This is surely one of the odder biological associations, that
of a one "unicellular" organism with other, symbiotic,
unicellular organisms living it. Zooxanthellate forams are
quite commonly found in natural reefs, but are less abundant
in our systems, possibly because the light levels are lower
than is normally found in many shallow water communities.
 |
Figure
6. Foram containing zooxanthellae collected from my aquarium.
Many forams are themselves food for other
aquarium denizens. Hermit crabs will eat them as will many
snails and some bristle worms. There are microscopic predators,
as well; some nematodes (roundworms) and flatworms will eat
them. The sand-sifting fishes are likely eating forams, among
other things, that they extract from the sand. Such predation
pressure, of course, will depress the foram populations, but
is not likely to have significant effects as long as there
are not too many predators. Like most amoebas, forams may
reproduce by binary fission, or splitting into two. Forams,
however, may at other times reproduce sexually producing many
hundreds of offspring. If conditions are good, they will rapidly
spread throughout the sand bed.
Conclusion
Beauty is not limited to larger things.
On a microscopic scale, foraminiferans often possess a sculptured
symmetry and architecture that is incomparable. They are fascinating,
if minute, organisms found in many of our aquaria. Their small
size belies their importance at helping us with our aquarium
maintenance. As with many of the other small organisms found
in sand beds, many of the larger, more decorative of our reef
aquarium species would simply be impossible to keep without
the contribution of foraminiferans. If you want to learn more
about foraminiferans, I would suggest you check out invertebrate
zoology textbooks at your local library, or do online searches
using the following terms: foraminifera, or foraminiferida.
There is a wealth of information on this ecologically very
important group.
|