|
Regular readers of this column, provided
there are regular readers of this column, will realize
that I like animals that tend to be odd, unusual, or simply
bizarre. The animals in the group I will write about this
month, the ctenophores,
(pronounced "teen'-oh-fores") really satisfy all
of those criteria. Interestingly enough, it is also a group
that I thought I would never be writing about when I started
commenting on the invertebrates found in reef aquaria. The
reason for that viewpoint is pretty obvious, the vast majority
of ctenophores
are wholly planktonic and, generally, as hobbyists, we simply
can't maintain truly planktonic invertebrate animals for any
length of time in any sort of normal condition. However, in
my blithe ignorance of what might turn up in tanks from time-to-time,
I neglected to realize that representatives from several species
of the relatively uncommon benthic ctenophores would hitchhike
their way into our systems with some regularity. Over the
past couple of years, it has become apparent that not only
are the crawling ctenophores present in some systems, they
are reproducing and living
quite successfully in them. Additionally, they are quite
widespread through the hobby.
Ctenophores
are some of the most common planktonic animals in many oceanic
realms, and in a discussion of their natural history it is
easier to work, at least initially, with some of the common
planktonic forms rather than the sessile ones. This is because
the planktonic forms are largely transparent, and it is relatively
easy to see most anatomical structures just by looking through
them. The crawling forms are often opaque and quite highly
colored. Additionally, while the sessile forms are very highly
modified and changed from their planktonic ancestors, they
do retain many basic features of those ancestors so that a
knowledge of the basic planktonic form helps significantly
in understanding the bottom crawlers.
|
Figure 1. A pelagic ctenophore, Pleurobrachia
bachei, photographed in nature showing some of the
morphology. Image copyright © Dr. Ronald L. Shimek,
2004.
|
These animals are placed by taxonomists
in the Phylum Ctenophora. This is one of the smaller phyla,
as there are only an estimated eighty to one hundred species.
They are all marine; there are no freshwater or terrestrial
species. They bear a superficial resemblance to some of the
various jellyfishes which are life stages of some of the cnidarians,
but they were recognized as being in a separate phylum by
Hatschek in 1889. Nonetheless, some textbooks continued to
treat them as part of a larger grouping called the coelenterates
until the late twentieth century. The so-called coelenterata
was an inappropriate combination of animal groups, containing
all of the cnidaria (animals such as hydroids, corals, and
jellyfishes) as well as the ctenophorans. This grouping was
based on a superficial similarity of form between ctenophorans
and some jellyfishes, coupled with the observation that one
ctenophoran species, Haeckelia
rubra, uses nematocysts to catch its prey. As nematocysts
are really a unique property of the cnidaria (no other group
has anything like them), this usage by the one ctenophoran
was deemed sufficient to allow the combining of the two groups.
Unfortunately for this view of the gelatinous zooplankton,
we now know that no ctenophore secretes nematocysts. Haeckelia
rubra, however, does use them. It eats medusae, and obtains
the nematocysts from one prey animal to assist in capturing
others (See Mills and Miller, 1984).
Structural Properties:
These are moderately-sized animals; the
body ranges from 5mm to about 1 m in length, depending on
the species. The tentacles of some of the larger species can
easily extend about 20 m or more. They are rather simply organized;
most of their structures are comprised of tissues; they have
relatively few structures that could be called organs. Primitive
forms have what may be called biradial symmetry. In other
words, while most structures are arranged in a radial pattern
around the axis of symmetry, many of these structures are
arranged in a mirror image pattern on either side of the central
plane of symmetry. Most pelagic ctenophores are roughly cylindrical
with discrete "lateral" sides. However, they don't
have a top or bottom, and may rotate around the center axis
as the animal swims. Consequently, unlike a fish, pelagic
ctenophores are not considered to be bilaterally symmetrical.
The tendency toward becoming a fully bilateral animal with
a front and back end as well as left and right sides reaches
its acme in this group in the benthic, crawling species. These
animals are quite "flatwormy" in appearance and
are really bilaterally symmetrical.
As with most radially, and biradially symmetrical
animals, they have an oral-aboral axis of symmetry. In other
words, the gut runs right down the centerline of the more
primitive forms; but they lack a brain and anything that might
be considered to be a head. They do have a sensory region
that surrounds, or is associated with, one end of the gut
tube. This is pretty promising for armchair classifiers. It
seems to indicate that ctenophores are really related to the
mainstream of invertebrate evolution. This is because a neural
aggregation with a sensory region surrounding the gut is really
"THE" invertebrate way to make an animal. In annelids,
mollusks, arthropods and an impressive array of smaller critters,
the brain or its components surround the gut in the region
of the esophagus or throat. In a snail, an octopus, or a shrimp,
it is no exaggeration to say that the gut passes right through
the brain. It would be tempting to say that ctenophores have
something similar, except that their sensory nervous aggregation,
called the polar field, is at the posterior end of the animal
near the anal pores (they don't have just one anus either,
but we will get to that momentarily).
 |
|
Figure
2. This image, taken from Figure 209, Hyman, L.
H. 1940, The Invertebrates, Protozoa through Ctenophora,
Volume 1. McGraw-Hill Book Company. New York, shows
the typical illustration of an inverted ctenophore.
Hyman wrote a six volume series considered to be the
classic English-language invertebrate reference and
certainly knew the proper orientation of the animals.
Note the inverted body and trailing tentacular fringe
of the ctenophore implying that the animal moved upward.
|
Biologists are a rather conservative breed
of human, and as a group, they generally consider it to be
dogma that an animal's brain must be near its front end. Simple
animals often lack a well-defined brain, but they often have
sensory structures near the front end of the body. These are
generally considered to be "evolutionary" precursors
of brains. Well, this neural and sensory aggregation of the
ctenophoran rear-end has really caused some of these armchair
biologist theoreticians to have an indigestible case of circular
reasoning.
It goes like this
- Brains are found near the front end
of animals.
- The ctenophoran sensory field and neural
aggregation is a primitive brain.
- Thus, the sensory field and neural aggregation
is at the front end of the animal.
- If you look through old editions of
many textbooks, and a few new editions as well, you will
find that the diagrams of the typical ctenophores show
the
sensory region at the top or at the presumed front
of the animal where a normal animal would have its head.
The mouth is drawn pointing down or to the rear.
Uh-huh…
These animals move mouth first, just like
most good bilateral animals do through the rest of the animal
kingdom. It would be ridiculous to say they move backwards
through the world, but that is just exactly what has been
assumed. Such a simple and "tidy" assumption about
these beautiful animals; it makes everything so nice, keeping
all presumptive brains up front. Well, that is all well and
good, but simple observation shows that the animals move mouth
first, and if the anally located polar sensory field helps
the animal orient (which it seems to), then we can truly say
that these animals are being given a bum steer… or, at
least, steering directions from their bum.
With few exceptions, ctenophores have a
pair of very extensible tentacles. The position of the tentacles
on each side of the body is what gives most ctenophores their
basic appearance of having two sides rather than being completely
radially symmetrical. Internally, the gut also is divided
into two halves as well, but that is not seen without magnification.
The tentacles are branched, and the branches extend from the
main tentacle axis like a fringe on only one side. They are
made of a muscular central core that is surrounded by a layer
of epidermis containing colloblast cells. Colloblasts are
"glue" cells and there are several different types
of them. When the tentacle contacts a prey item, the colloblasts
explode releasing adhesive strands and granules and these
adhere the tentacle to the prey. The prey, typically a small
crustacean, struggles when the tentacles start to stick to
them and this results in more of the tentacle getting wrapped
around the prey item. The ctenophore then swims so that the
mouth contacts the tentacle and eats the prey. Unlike the
discharging of cnidarian nematocysts which are non-living,
the act of capturing food by a ctenophore results from the
destruction of many cells in the tentacular epidermis.
 |
|
Figure
3. A smear preparation of a ctenophore tentacle
stained with Methylene Blue, photographed using a microscope
at 100x. There are many colloblasts in the field of
view; one is labeled to show some of the parts. Image
copyright © Dr. Ronald L. Shimek, 2004.
|
Unlike the bodies of jellyfish or most
other gelatinous planktonic animals, ctenophore bodies are
relatively rigid. They do not move by muscular means, which
is yet another difference between ctenophores and medusa.
Instead of propulsion by muscular contraction, they move by
paddling their way through the world. They have eight rows
of small paddles running along the body. In the primitive
forms these rows are evenly spaced, but in some others, such
as the benthic ones found in aquaria, the rows have been displaced.
The paddles are called "ctenes," a word meaning
"combs." They look quite like a miniature version
of a styling comb used to keep a hairdo in place. Each of
these combs is formed by several hundred large cilia which
have been fused together in a common sheath. They are arranged
in rows, called, not surprisingly, ctene rows, and the beat
of these is controlled by the sensory polar field.
The aboral sensory region or polar field
contains numerous sensory components such as statocysts which
give information about orientation and which are connected
directly to nerves that coordinate the locomotory beat of
the ctenes. Additionally, the sensory region also contains
photoreceptors or eyespots. Ctenophores do not respond directly
to shadows and the eyespots can't form an image. However,
they probably serve to keep track of changing day length and
may indicate to the animal if it has descended too far down
into the darkened depths. The sensory area also contains cells
that have stiff cilia that project into the water. These cilia
bend under the force of water moving past them, and probably
indicate to the animal how fast it is moving. Ctenophores
don't have nerves as we know them from vertebrates. In animals
such as us, a nerve is a collection of nerve fibers or nerve
cell processes which can extend some relatively great distance
from the cell body. The nerve cell bodies are not in the nerves
proper, but reside in the brain or in specialized aggregations
of nerve cell bodies called ganglia. In ctenophores, the nerves
are comprised of a mixture of cell bodies and relatively short
fibrillar processes.
The mouth is slitlike and found on the
end of the animal away from the sensory field. Typically,
the mouth is relatively small and oriented perpendicular to
the axis of the tentacles. Inside the animal is a rather capacious
gut region referred to as the stomodeum. At about the midpoint
of the gut, one branch arises from either side of the stomodeum.
These branches each subdivide two mores times to form eight
elongate gut pouches which are positioned under and parallel
to the ctene rows. Digestion starts in the stomodeum, but
most digestion is in the epithelia lining the gut directly
under the ctene rows. Nutrients are easily and rapidly transferred
to the ctene rows by diffusion, providing the animal with
food and energy to move about. Indigestible food remains are
passed out the anal pores at the aboral pole.
Reproduction and Development
Ctenophores are hermaphroditic, and the
gonads typically develop at separate times. The gonads exist
in discrete rows in the gut pouches under the ctene rows and
develop from the gut epithelium. Gametes escape via the anal
pore or through special gonoducts (found in only a few species).
Fertilization occurs in the sea and embryonic
development is very rapid. Generally, a characteristic
larva called a "cydippid" is found by the end of
the second day after spawning. Cydippids are small ctenophores
which have only four rather than eight ctene rows. If food
is abundant, these larvae will grow rapidly and may be reproductive
within a few more days. The most abundant ctenophore, the
sea gooseberry, Pleurobrachia bachei, may live a couple
of years. The potential life spans of most ctenophores are
unknown.
 |
 |
Figure 4. Left. A cydippid larva of a pelagic
ctenophore, probably Pleurobrachia bachei.
Right. A cydippid larva of a platyctene ctenophore
found in an aquarium. Note the similarities
of structure. Images copyright © Dr. Ronald L. Shimek,
2004.
There are several different taxonomic orders
of planktonic ctenophores, while only one order contains species
adapted to live on substrates. This latter group, called the
"Order Platyctenida," contains those ctenophores
found in marine reef aquaria. These animals have a flattened
body that looks very "flatwormish," but differs
from the flat worms in the presence of the ctene rows on the
ventral surface and a pair of perfectly normal ctenophore
tentacles that arise from pouches on the dorsal surface. Platyctenes
may be visualized as a "deflated" example of a more
typical pelagic ctenophore. The gut pouches lie internally
in groups of four on either side of the animal, and differential
growth has resulted in the ctene rows all being on the bottom
and the tentacles being on the top. Additionally, they are
colored and often have relatively ornate patterns on their
upper surfaces. This "ornamentation" often matches
the surfaces of the animals, such as the specific species
of soft corals, or sea
stars(1) (2),
they live on. The color pattern probably provides them with
some degree of protection from visual predators such as "nipping"
fishes. Other sessile ctenophores, such as this Antarctic
Lyrocteis, are neutrally colored.
 |
Figure 5. An unidentified Platyctene ctenophore found
in an aquarium. The tentacles are extruded but
not extended due to the lack of current in the bowl where
the photography was done.
Image copyright © Dr. Ronald L. Shimek, 2004.
Figure 6. An unidentified Platyctene ctenophore found
in an aquarium; possibly the same species as in
Figure 5, but collected and sent to me several months earlier.
The tentacles are retracted; note the
flatworm-like appearance. Image copyright © Dr. Ronald
L. Shimek, 2004.
Platyctenes are often found living on particular
soft corals or corals, and may also absorb or eat mucus from
their "host." It is unlikely that they actually
eat the host's tissues. Platyctenes, generally, do not seem
to specifically harm animals in reef aquaria, provided that
there are only a few of the ctenophores. In a few instances,
however, they have achieved plague proportions and may be
so abundant that they actually smother their hosts or other
animals. This later result is quite uncommon. In most tanks,
they enter as hitchhikers, persist for a few weeks and then
vanish.
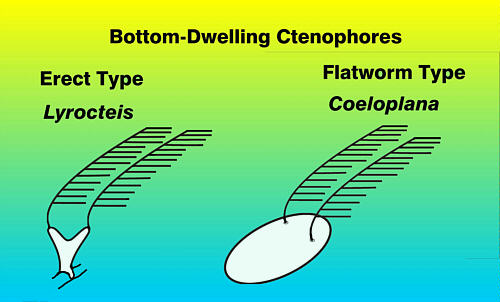
Figure 7. Diagrams of benthic ctenophores showing basic
shapes. So far, only the "flatworm"
type has been found in aquaria.
Although both pelagic and benthic ctenophores
have been kept for long times in research aquaria (I have
kept an individual of one pelagic species, Beroe, for
several months - but I had ready access to its prey, another
species of ctenophore, Pleurobrachia, to feed it),
pelagic species generally require the same special conditions
that medusae need. So, typically only large commercial aquaria,
such as the Monterey Bay Aquarium, can afford to produce and
maintain the large circular laminar flow tanks necessary so
that the animals do not continually bounce themselves off
the aquarium walls. I don't know of any hobbyist who has successfully
kept any of the swimming forms. However, I would expect that
sooner or later some will be kept. These are strikingly beautiful
animals; the beating ctene rows are almost always iridescent
and under normal lighting the animals give the appearance
of having flickering rainbows on each ctene row, and the body
shapes are intricate and strange, which adds to their fascination.
 |
Figure 8. Note the iridescence of the ctene rows on
this ctenophore,
Bolinopsis. Image copyright © Dr. Ronald L. Shimek,
2004.
Incidentally, exotic or introduced ctenophore
species have become problems in several regions of the world.
Although simple and relatively small, ctenophores are voracious
predators, and this fact, coupled with their astronomical
reproductive rate can result in wholesale changes to the organism
arrays if they are inadvertently introduced into environments
where they were previously not found. Sometimes these introductions
result in wholesale faunal changes. One such change has been
recently documented concerning a ctenophore, native to the
Atlantic, which has been introduced into the Black
Sea. Probably transported in a ship's ballast water, this
ctenophore, Mnemiopsis leidyi, is a common component
of the North Atlantic. It was first noted in the Black Sea
about 1986, and has had a devastating effect on the native
fauna and human fisheries. It has recently been found in the
Caspian
Sea, and will undoubtedly destroy the native ecosystems
in that region as well. Check out the link to the information
about this environmental disaster for some interesting data
on a very ugly event.
For a further detailed, but very readable,
discussion of Ctenophores, I recommend Dr.
Claudia Mills' website as a place to begin.
|