|
References:
1. The structure of sea urchin spines,
large biogenic single crystals of calcite. Su, X.; Kamat,
S.; Heuer, A. H. Department of Materials Science and Engineering,
Case Western Reserve University, Cleveland, OH, USA. Journal
of Materials Science (2000), 35(22), 5545-5551.
2. Single crystal structure analysis of sea urchin spine
calcites: systematic investigations of the Ca/Mg distribution
as a function of habitat of the sea urchin and the sample
location in the spine. Magdans, Uta; Gies, Hermann. Institut
GMG, Ruhr-Universitaet Bochum, Bochum, Germany. European Journal
of Mineralogy (2004), 16(2), 261-268.
3. Structure of magnesian calcite from sea urchins.
Tsipursky, Semeon J.; Buseck, Peter R. Dep. Geol., Arizona
State Univ., Tempe, AZ, USA. American Mineralogist (1993),
78(7-8), 775-81.
4. Accumulation of Pb and Zn in sea urchin plates and
spines related to their different crystalline structure.
Chinchon, S.; Auernheimer, C.; Alastuey, A.; Gali, S. Departamento
de Construcciones Arquitectonicas, Universidad de Alicante,
Alicante, Spain. Marine Pollution Bulletin (2000), 40(7),
647-649.
5. Allocation of calcium-45 to body components of starved
and fed purple sea urchins (Strongylocentrotus purpuratus).
Lewis, C. A.; Ebert, T. A.; Boren, M. E. Dep. Biol., San Diego
State Univ., San Diego, CA, USA. Marine Biology (Berlin, Germany)
(1990), 105(2), 213-22.
6. Does calcium carbonate in food deter feeding by sea
urchins? Pennings, Steven C.; Svedberg, Joan Marie. Mar.
Lab., Univ. Guam, Mangilao, Guam. Marine Ecology: Progress
Series (1993), 101(1-2), 163-7.
7. Secondary metabolites and calcium carbonate as defenses
of calcareous algae on coral reefs. Paul, V. J. Marine
Lab., Univ. Guam, Mangilao, Guam. Editor(s): Lessios, Harilaos
A.; Macintyre, Ian G. Proceedings of the International Coral
Reef Symposium, 8th, Panama, June 24-29, 1996 (1997), Meeting
Date 1996, 1 707-712.
8. Biomineralization of the spicules of sea urchin embryos.
Wilt, Fred H. Dept. of Molecular Cell Biology, University
of California, Berkeley, USA. Zoological Science (2002), 19(3),
253-261.
9. Interactions of sea-urchin skeleton macromolecules
with growing calcite crystals - a study of intracrystalline
proteins. Berman, A.; Addadi, L.; Weiner, S. Dep. Isot.
Res., Weizmann Inst. Sci., Rehovot, Israel. Nature (London,
United Kingdom) (1988), 331(6156), 546-8.
10. Interactions of various skeletal intracrystalline
components with calcite crystals. Albeck, S.; Aizenberg,
J.; Addadi, L.; Weiner, S. Dep. Struct. Biol., Weizmann Inst.
Sci., Rehovot, Israel. Journal of the American Chemical Society
(1993), 115(25), 11691-7.
11. Proteins and Saccharides of the Sea Urchin Organic
Matrix of Mineralization: Characterization and Localization
in the Spine Skeleton. Ameye, Laurent; De Becker, Genevieve;
Killian, Christopher; Wilt, Fred; Kemps, Raymond; Kuypers,
Stephan; Dubois, Philippe. Laboratoire de Biologie Marine,
Universite Libre de Bruxelles, Brussels, Belg. Journal of
Structural Biology (2001), 134(1), 56-66.
12. Polysaccharides of intracrystalline glycoproteins
modulate calcite crystal growth in vitro. Albeck,
Shira; Weiner, Steve; Addadi, Lia. Dep. Struct. Biol., Weizmann
Inst. Sci., Rehovot, Israel. Chemistry--A European Journal
(1996), 2(3), 278-84.
13. Regulation of calcite crystal morphology by intracrystalline
acidic proteins and glycoproteins. Albeck, S.; Addadi,
L.; Weiner, S. Dept. Structural Biology, Weizmann Institute
Science, Rehovot, Israel. Connective Tissue Research (1996),
35(1-4), 365-370.
14. Spicule matrix protein LSM34 is essential for biomineralization
of the sea urchin spicule. Peled-Kamar, Mira; Hamilton,
Patricia; Wilt, Fred H. Molecular Cell Biology Department,
University of California, Berkeley, CA, USA. Experimental
Cell Research (2002), 272(1), 56-61.
14. The transient phase of amorphous calcium carbonate
in sea urchin larval spicules: the involvement of proteins
and magnesium ions in its formation and stabilization.
Raz, Sefi; Hamilton, Patricia C.; Wilt, Fred H.; Weiner, Steve;
Addadi, Lia. Department of Structural Biology, Weizmann Institute
of Science, Rehovot, Israel. Advanced Functional Materials
(2003), 13(6), 480-486.
16. Superimposed effect of kinetics and echinoderm glycoproteins
on hierarchical growth of calcium carbonate. MacKenzie,
Callum R.; Wilbanks, Sigurd M.; McGrath, K. M. Department
of Chemistry, School of Chemical and Physical Science, Victoria
University of Wellington, Wellington, N. Z. Journal of Materials
Chemistry (2004), 14(8), 1238-1244.
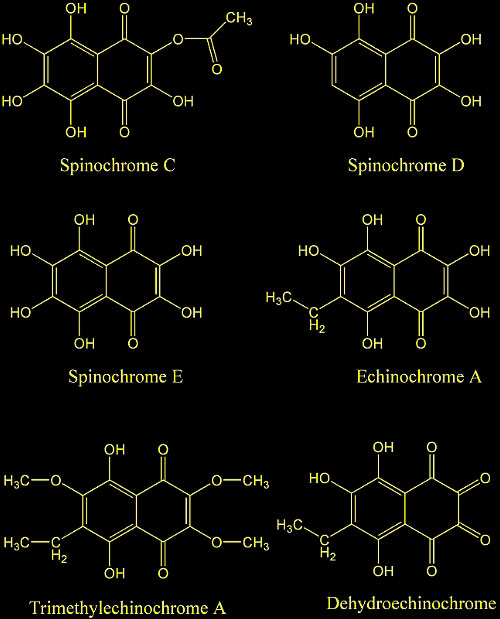
17. Sephadex LH-20 separation of pigments from shells
of red sea urchin (Strongylocentrotus franciscanus).
Amarowicz, R.; Synowiecki, J.; Shahidi, F. Department of Biochemistry,
Memorial University of Newfoundland, St. John's, NF, Can.
Food Chemistry (1994), 51(2), 227-9.
18. Quinoid pigments of echinoderms. VIII. Pigments of
sea urchins Diadema setosum and Diadema savignije.
Kol'tsova, E. A.; Maksimov, O. B. Tikhookean. Inst. Bioorg.
Khim., Vladivostok, USSR. Khimiya Prirodnykh Soedinenii (1981),
(1), 115.
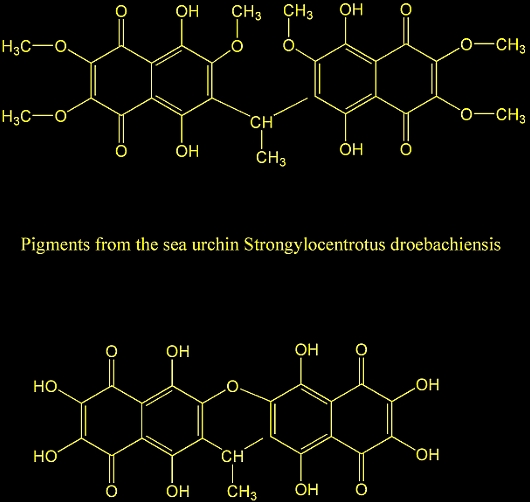
19. Quinoid pigments from echinoderms. V. Pigments from
the sea urchin Strongylocentrotus droebachiensis.
Kol'tsova, E. A.; Denisenko, V. A.; Maksimov, O. B. Dal'nevost.
Nauch. Tsentra, Tikhookean. Inst. Bioorg. Khim., Vladivostok,
USSR. Khimiya Prirodnykh Soedinenii (1978), (4), 438-41.
20. Quinoid pigments of echinoderms. III. Minor pigments
of the sea urchin Strongylocentrotus nudus. Kol'tsova,
E. A.; Chumak, G. N.; Maksimov, O. B. Tikhookean. Inst. Bioorg.
Khim., Vladivostok, USSR. Khimiya Prirodnykh Soedinenii (1977),
(2), 202-7.
21. Derivatives of naphthoquinones. XIII. Pigments from
sea urchins. 8. Kuroda, Chika; Harada, Miye. Sci. Research
Inst., Tokyo, Proc. Japan Acad. (1955), 31 305-8.
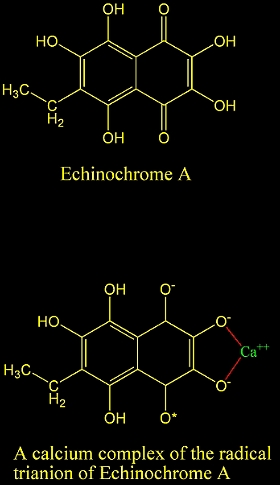
22. How do calcium ions induce free radical oxidation
of hydroxy-1,4-naphthoquinone? Ca2+ stabilizes the naphthosemiquinone
anion-radical of echinochrome A. Lebedev, Alexander V.;
Ivanova, Marina V.; Ruuge, Enno K. Institute of Experimental
Cardiology, Department of Biochemistry, Cardiology Research
Center, Moscow, Russia. Archives of Biochemistry and Biophysics
(2003), 413(2), 191-198.
23. Antifouling agent for fish net. Ashino, Izumi.
(Andrews and George Co., Inc., Japan). Jpn. Kokai Tokkyo Koho
(1977), 2 pp.
24. Biocidal compositions comprising polyhydroxynaphthoquinones.
Sime, Alan Alexander Torrance. (UK). Brit. UK Pat. Appl. (1985),
11 pp.
25. Biocidal compds. for controlling populations of aquatic
pest organisms containing quinones, anthraquinones, and naphthalenediones.
Cutler, Horace; Cutler, Stephen; Wright, David; Dawson, Rodger.
(Garnett, Inc., USA). PCT Int. Appl. (2001), 30 pp.
26. Antimicrobial activity of polyhydroxynaphthoquinones
from sea urchins. Stekhova S I; Shentsova E B; Kol'tsova
E B; Kulesh N I Antibiotiki i khimioterapiia = Antibiotics
and chemoterapy [sic] / Ministerstvo meditsinskoi i mikrobiologicheskoi
promyshlennosti SSSR (1988 Nov), 33(11), 831-3.
27. Chemistry of Superoxide Radical in Seawater: CDOM
Associated Sink of Superoxide in Coastal Waters. Goldstone,
Jared V.; Voelker, Bettina M. Department of Civil and Environmental
Engineering, Massachusetts Institute of Technology, Cambridge,
MA, USA. Environmental Science and Technology (2000), 34(6),
1043-1048.
28. Chemistry of Superoxide Radical in Seawater: Reactions
with Organic Cu Complexes. Voelker, Bettina M.; Sedlak,
David L.; Zafiriou, Oliver C. Department of Marine Chemistry
and Geochemistry, Woods Hole Oceanographic Institution, Woods
Hole, MA, USA. Environmental Science and Technology (2000),
34(6), 1036-1042.
29. Fate of superoxide in coastal sea water. Petasne,
Robert G.; Zika, Rod G. Rosenstiel Sch. Mar. Atmos. Sci.,
Univ. Miami, Miami, FL, USA. Nature (London, United Kingdom)
(1987), 325(6104), 516-18.
30. Chemistry of the superoxide radical (O2-) in seawater:
Reactions with inorganic copper complexes. Zafiriou, Oliver
C.; Voelker, Bettina M.; Sedlak, David L. Woods Hole Oceanographic
Institution, Woods Hole, MA, USA. Journal of Physical Chemistry
A (1998), 102(28), 5693-5700.
31. Superoxide radical production by sponges Sycon
sp. Peskin, Alexander V.; Labas, Yulii A.; Tikhonov, Alexander
N. Institute of Developmental Biology, Russian Academy of
Sciences, Moscow, Russia. FEBS Letters (1998), 434(1,2), 201-204.
32. Interaction of natural polyhydroxy-1,4-naphthoquinones
with superoxide anion-radical. Lebedev, A. V.; Ivanova,
M. V.; Krasnovid, N. I. Institute of Experimental Cardiology,
Cardiology Research Center, Russian Ministry of Health, Moscow,
Russia. Biochemistry (Moscow)(Translation of Biokhimiya (Moscow))
(1999), 64(11), 1273-1278.
33. Mechanisms of inhibition of iron(2+)-induced oxidation
of phosphatidylcholine by polyhydroxynaphthoquinones.
Lebedev, A. V.; Boguslavskaya, L. V.; Levitskii, D. O.; Maksimov,
O. B. Inst. Exp. Cardiol., Cardiol. Res. Cent., Moscow, USSR.
Biokhimiya (Moscow) (1988), 53(4), 598-603.
34. Exposure to ultraviolet radiation causes apoptosis
in developing sea urchin embryos. Lesser Michael P; Kruse
Valerie A; Barry Thomas M Department of Zoology and Center
for Marine Biology, University of New Hampshire, Durham, NH
03824, USA. mpl@cisunix.unh.edu Journal of experimental biology
(2003 Nov), 206(Pt 22), 4097-103.
35. Synchrotron X-ray studies of the keel of the short-spined
sea urchin Lytechinus variegatus: absorption microtomography
(microCT) and small beam diffraction mapping. Stock S
R; Barss J; Dahl T; Veis A; Almer J D; Carlo F Institute for
Bioengineering and Nanoscience in Advanced Medicine, Northwestern
University, Chicago, IL 60611, USA. s-stock@northwestern.edu
Calcified tissue international (2003 May), 72(5), 555-66.
36. Mineral phase microstructure in teeth of the short
spined sea urchin (Lytechinus variegatus) studied with
X-ray phase contrast imaging and with absorption microtomography.
Stock, S. R.; Dahl, Thomas; Barss, Joseph; Veis, Arthur; Fezzaa,
K.; Lee, W. K. School of Materials Sci. & Eng., Georgia
Inst. of Technology, Atlanta, GA, USA. Advances in X-Ray Analysis
(2002), Volume Date 2001, 45 133-138.
37. Mineral-related proteins of sea urchin teeth: Lytechinus
variegatus. Veis Arthur; Barss Joseph; Dahl Thomas;
Rahima Mohammed; Stock Stuart Department of Cell and Molecular
Biology, Northwestern University Medical School, Chicago,
Illinois 60611, USA. aveis@northwestern.edu Microscopy research
and technique (2002 Dec 1), 59(5), 342-51.
38. Design strategies of sea urchin teeth: structure,
composition and micromechanical relations to function.
Wang R Z; Addadi L; Weiner S Department of Structural Biology,
Weizmann Institute of Science, Rehorol, Israel Philosophical
transactions of the Royal Society of London. Series B, Biological
sciences (1997 Apr 29), 352(1352), 469-80.
39. Matrix proteins of the teeth of the sea urchin Lytechinus
variegatus. Veis D J; Albinger T M; Clohisy J; Rahima
M; Sabsay B; Veis A Journal of experimental zoology (1986
Oct), 240(1), 35-46.
40. Natural carbonate-reinforced composite materials.
Part 1. Morphology of sea urchin teeth. Bonfield, W.;
Scandola, M. Dep. Mater., Queen Mary Coll., London, UK. Journal
of Materials Science (1979), 14(12), 2865-71.
41. X-ray absorption microtomography (microCT) and small
beam diffraction mapping of sea urchin teeth. Stock S
R; Barss J; Dahl T; Veis A; Almer J D Institute for Bioengineering
and Nanoscience in Advanced Medicine, Northwestern University,
Chicago, IL 60611-3008, USA. s-stock@northwestern.edu Journal
of structural biology (2002 Jul), 139(1), 1-12.
42. Recent Studies on the Pathological Effects of Purified
Sea Urchin Toxins. Nakagawa, H.; Tanigawa, T.; Tomita,
K.; Tomihara, Y.; Araki, Y.; Tachikawa, E. Department of Life
Sciences, University of Tokushima, Tokushima, Japan. Journal
of Toxicology, Toxin Reviews (2003), 22(4), 633-649.
43. The inhibitory effect of the toxic fraction from sea
urchin (Toxopneustes pileolus) venom on 45Ca2+ uptake
in crude synaptosome fraction from chick brain. Zhang,
Yu-An; Wada, Tetsuyuki; Iwasaki, Yasunori; Nakagawa, Hideyuki;
Ichida, Seiji. Department of Biological Chemistry, Faculty
of Pharmaceutical Sciences, Kinki University, Higashi-Osaka,
Japan. Biological & Pharmaceutical Bulletin (1999), 22(12),
1279-1283.
44. Mode of attack of sea urchin toxin on natural and
synthetic substrates. II. Physical properties, substrate specificity,
and reaction kinetics of purified fractions. Feigen, George
A.; Hadji, Lahlou; Pfeffer, Roger A.; Markus, Gabor. Dep.
Physiol., Stanford Univ., Stanford, CA, USA. Physiological
Chemistry and Physics (1970), 2(5), 427-44.
45. Fishing for bioactive substances from scorpionfish
and some sea urchins. Satoh, F.; Nakagawa, H.; Yamada,
H.; Nagasaka, K.; Nagasaka, T.; Araki, Y.; Tomihara, Y.; Nozaki,
M.; Sakuraba, H.; Ohshima, T.; Hatakeyama, T.; Aoyagi, H.
Department of Life Sciences, University of Tokushima, Tokushima,
Japan. Journal of Natural Toxins (2002), 11(4), 297-304.
46. Purification and characterization of contractin A
from the pedicellarial venom of sea urchin, Toxopneustes
pileolus. Nakagawa, Hideyuki; Tu, Anthony T.; Kimura,
Akira. Dep. Biochem., Colorado State Univ., Fort Collins,
CO, USA. Archives of Biochemistry and Biophysics (1991), 284(2),
279-84.
47. Effect of Contractin A, a glycoprotein from the pedicellarial
venom of the sea urchin Toxopneustes pileolus, on isolated
vascular smooth muscles. Nakagawa, Hideyuki; Kitagawa,
Hisato; Kondo, Noriaki; Kondo, Jun. Fac. Integrated Arts Sci.,
Univ. Tokushima, Tokushima, Japan. Journal of Natural Toxins
(1992), 1(2), 31-7. CODEN: JNTOER ISSN: 1058-8108.
48. Preliminary studies on venom proteins in the pedicellariae
of the toxopneustid sea urchins, Toxopneustes pileolus
and Tripneustes gratilla. Nakagawa, Hideyuki; Tomihara,
Yasuhiro; Araki, Yasutetsu; Hayashi, Hiromi. Dep. Life Sci.,
Univ. Tokushima, Tokushima, Japan. Journal of Natural Toxins
(1994), 3(1), 25-34.
|